In these days of such uncertainty, not all the news is bad with the Coronavirus. Scientists in China are starting to test regenerative medicine treatments to reduce this pandemic that affects the whole world. Dr. Dongcheng has already treated 9 patients who had been hospitalized with the virus with satisfactory results. The idea behind the treatment is to reduce the level of pneumonia and inflammation in the lungs through IV infusion of cord-derived allogeneic stem cells.
This type of therapy has been performed mainly in COPD patients with the aim of reducing inflammation in the lungs. Normally when stem cells are given intravenously, they pass through a pulmonary pathway that essentially captures the stem cells where they are needed most in the patient’s body. Those stem cells and growth factors can then be effective in reducing inflammation within the lungs.
Several studies have shown that mesenchymal stem cells can differentiate into several different types of lung cells, such as alveolar epithelial cells, which are destroyed by various chronic conditions that cause emphysemic changes and reduced support of the small airways, causing hyperinflation and gas exchange abnormalities. Pre-clinical trials have suggested regeneration of alveolar structures, repair of emphysemic lungs and reduction of inflammatory responses.
There are currently several doctors and companies working with the FDA and CDC in the United States to begin using stem cells. Even companies like Cord for Life promise 600K in products if the treatment gets approved. But the reality is that if it is true that cells could be an effective therapy for COPD, so far we lack sufficient data to claim that against coronavirus. However, we believe that it is worth trying, we are in a moment of crisis, people are dying and we cannot wait months for this therapy to be approved by the FDA.
Being a doctor is unarguable, one
of the most highly regarded and prestigious profession. There are several
mistakes doctors make without realizing it. As a physician, several qualities
make you stand out and give you an edge over others. Have you tried to put yourself in the shoes
of your patients? If you have, you will realize that there are several things
expected of you as Doctor, which you may or may not be doing well.
As a physician, you need to have
encyclopedic medical knowledge, especially those in the regenerative medicine
field. Having a comprehensive knowledge is not all about what you know at the
moment but about what you will learn every day and the in formations that will
keep you at the top in your field. With the amount of research being published
every day in medical news and trends, you need to be up-to-date because some
new information will contradict what was learned back in medical school.
Medicine is always changing, and there is still room for learning new
techniques and procedures that can benefit your patients and practice.
In our organization, one of our
big goals since we started, is providing physicians with the latest knowledge
about the therapeutic benefits available in regenerative medicine which can be
implemented in a medical office setting. We are focused on translational
medicine, that means we take a detail of every procedure happening in the
laboratories, and we make it available for doctors patients.
As a physician, your approach to
cases influences your patient’s response and how it impacts them. Most patients
would prefer a wrong diagnosis to be delivered at least with a hint of hope.
Your job as a physician is to give them a hint a hope not by giving them a
false diagnosis but being honest and providing accurate information but without
bluntness or factual information. You need to inspire your patients with
optimism because optimistic patients have better health outcome, even if you
become unsure about something, don’t let you patient see it. Patients want to
trust their physicians, listen to everything he/she says without preemptively
voicing your diagnosis.
Here in ISSCA (International
Society for Stem Cell Application), we provide certification and standard in
the practice of medicine. Our goal is to use regenerative medicine to treat
diseases and lessen human suffering.
In the medical community, every
manufacturer is talking endlessly about the advantage and disadvantage of using
cellular products. People are also more interested in medical treatments that
are more natural. All patients are eager to know what is available there that
could help them and relief their ailment because regenerative medicine and
cellular therapy have proven to offer so much hope. For quite a while now,
conventional treatments only target the symptoms and not the causes, unlike
cellular therapy.
As stem cell therapy is a
relatively new science, we are here to light the way for our members, thereby
giving them an edge in Regenerative medicines and cellular therapy.
ISSCA is set out to ensure you
find relief using regenerative medicine, with our endless research to ensure
patients get the best out of Cellular therapy. We are also focused on equipping
physicians, scientist and veterinarians who dedicated their quota to their
practice on regenerative medicine and cellular therapy and using this technique
to treat many disease conditions and improve health for a variety of diseases
and health condition.
Some people believe that stem cell does all of this but studies have shown that It’s unlikely only in a few unique cases. You can observe minimal growth a year after the patient took treatment, but this doesn’t mean replacement of the cartilage. The cartilage has a reduced regenerative capacity, and current and present pharmacological medications only offer symptomatic pain relief. Osteoarthritis patients that respond poorly to conventional therapies are ultimately treated with surgical procedures to promote cartilage repair by implantation of artificial joint structures (arthroplasty) or total joint replacement (TJR). Surgery has been the last resort for serious cartilage problems. In the last two decades, stem cells derived from various tissues with varying differentiation and tissue regeneration potential have been used for the treatment of osteoarthritis, damage to bones and others either alone or in combination with natural or synthetic scaffolds. The stem cells derived from these tissues primarily aid cartilage repair. Although stem cells can be differentiated into chondrocytes in vitro or aid cartilage regeneration in vivo, their potential for Osteoarthritis management remains limited as cartilage regenerated by stem cells fails to fully recapitulate the structural and biomechanical properties of the native tissue. It isn’t easy for the cartilage to regrow and assume its original biomechanical and structure form. Apparently, Due to the limited intrinsic capacity of resident chondrocytes to regrow the lost cartilage post-injury, stem cell-based therapies have been proposed as a novel therapeutic approach for cartilage repair. Also, stem cell-based therapies using mesenchyme stem cells (MSCs) or induced pluripotent stem cells (iPSCs) have been used successfully in clinical and preclinical situations. Part of the issues associated with Mesenchyme stem cells can be averted by using iPSCs. iPSCs are an ideal patient-specific unlimited cell source for autologous tissue regeneration. With the Promising in vitro; studies have shown that vitro results have already been demonstrated in the cartilage engineering field for iPSCs. These were generated from various cell types. What Is Cartilage and How Does It Get Damaged? Cartilage is a connective tissue in the human body and body of other animals. In our joints, we have a few kinds of cartilage, but most often people refer to the smooth lining of a joint called articular or hyaline cartilage. This kind of cartilage gives rise to a soft layer of cushion on the end of a bone at the joint. The cushion is essential for balance, mechanical functions and athletics. This tissue of the cartilage is very strong, yet it can compress, readjust and absorb varying degrees of energy. It is also very slippery, smooth and flexible and these features allow the joint to glide effortlessly through a broad range of physical motions of any kind. When joint cartilage is not working correctly or damaged, this smooth-cushioning-layer can be worn away, and this becomes a problem. In the case of traumatic injuries, sometimes a sudden force causes the cartilage to break off or poorly become damaged, exposing the underlying bone of the body. In the case of osteoarthritis (also called degenerative/wear-and-tear arthritis), over time that smooth layer can wear thin and uneven. Aging can also cause the cartilage to break off and certain life factors and diseases too, e.g. autoimmune diseases. Eventually, as that cushion of the bones wears away, joint movements can become inflexible, stiff and painful on one or both legs (bones). Joints can even become inflamed and swollen. And as all these conditions, typically causes pain and limitations in activity become problematic. The action or activities that involve these bones leads to crushing pain and discomfort, depending on the severity of the case. Almost all activities involve the movement of bones; hence this condition is not an easy one. There are some treatments for cartilage damage and arthritis. Although there some medicines, most of these treatments are focused either on relieving symptoms by smoothing down the damaged cartilage or concentrate on replacing the joint surface with an artificial implant. The later is for end-stage conditions, and the artificial plane is procedures such as knee replacement or hip replacement surgery. How Can Stem Cells Help? Stem cells are specialized cells that can multiply reform and develop into different types of tissue. In the developmental stages of a fetus, stem cells are plentiful and surplus. However, in adulthood, stem cells are restricted to specific tasks of regenerating a few types of cells, such as blood cells and liver cells in some cases of damage. There are almost no stem cells found in cartilage tissue, and therefore there is little to no capacity to heal or regrow new cartilage. For adults, the ability to regrow new cartilage is even more difficult due to age and lack of stem cells in the cartilages. Most often, in the setting of orthopedic surgery and joint problems, stem cells are obtained from adult stem cell sources. The primary sources are bone marrow and fatty tissue. These stem cells can develop into cartilage cells, called chondrocytes. They also exhibit some other helpful qualities by stimulating the body to reduce inflammation, stimulate cell repair, and improve blood flow. This process is caused by the secretion of cellular signals and growth factors to stimulate the body to initiate healing processes. Once stem cells have been obtained, they need to be delivered to the area of the cartilage that damaged. One option is to inject the stem cells into the joint. There have been many studies investigating just this, and some data shows improvement in symptoms. How much of this improvement is the result of new cartilage growth versus other effects of stem cells (the healing properties listed above, including the anti-inflammatory effects) is unknown. There is a challenge with giving stem cell injection. The problem with just injecting stem cells is that cartilage is a complex tissue that is comprised of more than only cells hence this can pose a challenge because the stem can’t regenerate all the things in the cartilage. To regrow the cartilage, the complex tissue structure and biomechanics of cartilage must also be reconstructed to its former status. Cartilage can often /described as having a scaffold-like structure that is composed of water, cells, collagen, and proteoglycans, and infection-fighting antibodies. Injecting just the stem cells is thought to be less effective in stimulating the formation of the entire cartilage structure hence the challenge. Some studies are investigating the types of 3-dimensional tissue scaffolds engineered to have a cartilage-like structure. The stem can then be injected into the scaffold, in hopes of better restoring a healthy type of cartilage. Three-dimensional printing is becoming an exciting part of this type of research. If everything works out as expected, the cartilage reconstruction could be achieved to a very high percentage. How do stem cells work? Necessarily, stem cells are progenitor cells which are capable of regeneration and differentiation into a wide range of specialized cell types. Once injected, stem cells follow inflammatory signals from damaged tissues and have multiple ways of repairing these damaged areas. It works as though the part is developing new; like what is seen during a child’s development. The mesenchyme stem cells (MSCs) we are using are considered to be multipotent (they can transform into different cell types but cannot form an organ) but not pluripotent. In the body, these cells Do NOT function by transforming into different cell types or tissues. They act via anti-inflammatory activity, immune modulating capacity, and the ability to stimulate regeneration. We go through a very high thorough screening process to find cells that we know have the best anti-inflammatory activity, the best immune modulating capacity, and the best ability to stimulate regeneration process on the tissue with damage. ISSCA (International Society for Stem Cells Applications) www.issca.us This is a business located in Miami, FL, where people around the world come to take a certification in the newest Stem Cells Protocols. Some organizations have put in efforts to help discover some solutions in stem medicine. International Society for Stem Cell Application (ISSCA ) is one of the leading associations in setting standards and promoting excellence in the field of Regenerative Medicine, researches, publications related education, certification, research and publications. The ISSCA is a unique-multidisciplinary community of physicians, stem specialist and scientists with a mission to advance the science, technology and practice of Regenerative Medicine. Their aim is to treat disease and lessen human suffering. ISSCA generally advances the specialty of Regenerative Medicine and serves its members. The ISSCA provides certifications and standards in the practice of Regenerative Medicine as a medical specialty. Although the expectation on this stem cell course is yet to be achieved; however, this is a part of medicine that can offer one-end-solution to various bone and body problems. With the recent high-tech studies, efforts and dynamics, stem cell treatment can be a breakthrough in the future as its perspectives are very promising and unique. It is also not dangerous on the long-run.
Induced pluripotent stem cells (also known as iPS cells or iPSCs) are a type of pluripotent stem cell that can be generated directly from a somatic cell. Pluripotent stem cells hold promise in the field of regenerative medicine. Because they can propagate indefinitely, as well as give rise to every other cell type in the body (such as neurons, heart, pancreatic, and liver cells), they represent a single source of cells that could be used to replace those lost to damage or disease.
Natural killer cells are the type of cytotoxic lymphocyte critical to the innate immune system. The role NK cells play is analogous to that of cytotoxic T cells in the vertebrate adaptive immune response. NK cells provide rapid responses to virus-infected cells, acting at around three days after infection, and respond to tumor formation.
Typically, immune cells detect the major histocompatibility complex (MHC) presented on infected cell surfaces, triggering cytokine release, causing apoptosis. NK cells are unique, however, as they can recognize stressed cells in the absence of antibodies and MHC, allowing for a much faster immune reaction.
Clinical Trial on NK cells
In a first clinical trial, a natural killer cell immunotherapy derived from induced pluripotent stem cells is being tested for safety in 64 patients with a variety of solid tumors. The first subjects used for the study received the cells in February at the University of California, San Diego (UCSD) Moores Cancer Center and MD Anderson Cancer Center.
This study is targeting late-stage cancer patients with solid tumors, including lymphoma, colorectal cancer, and breast cancer. The FT500 NK cells do not undergo any further alterations and after their derivation from the induced pluripotent stem cells (iPSCs), offering the possibility of a quicker, ready-made treatment.
Human embryonic stem cells induced iPSCs
Human embryonic stem cells (hESCs) and induced pluripotent stem cells (iPSCs) provide an accessible, genetically tractable, and homogenous starting cell population to efficiently study human blood cell development. These cell populations provide platforms to develop new cell-based therapies to treat both malignant and nonmalignant hematological diseases.
The NK cells are immune cells in the same family as T and B cells and are very good at targeting cancer cells for destruction. Some Laboratory experiments have shown they do so by attacking cells that have lost their significant self-recognition signals that tell the immune system not to attack. This is the phenomenon that can happen among cancer cells but not to healthy cells. Experts are not sure how many cancer cells lose that signal. Researchers are hopeful that the clinical trial can help determine which cancer patients could benefit the most from NK cell treatment.
iPS Clone
The ability to induce pluripotent stem cells from committed, human somatic cells provides tremendous potential for regenerative medicine. However, there is a defined neoplastic potential inherent to such reprogramming that must be understood and may offer a model for critical understanding events in the formation of the tumor. Using genome-wide assays, we identify cancer-related epigenetic abnormalities that arise early during reprogramming and persist in induced pluripotent stem cell (iPS) clones. These include hundreds of abnormal gene silencing events, patterns of aberrant responses to epigenetic-modifying drugs resembling those for cancer cells, and presence in iPS and partially reprogrammed cells of cancer-specific gene promoter DNA methylation alterations.
Progress in adoptive T-cell therapy for cancer and infectious diseases is hampered by the lack of readily available antigen-specific, human T lymphocytes. Pluripotent stem cells could provide an estimable source of T lymphocytes, but the therapeutic potential of human pluripotent stem cell-derived lymphoid cells generated to date remains uncertain.
Modification of T cells
Recently, some Approved cell therapies for Cancer also rely on modifying T cells, in those cases to produce cancer cell–binding chimeric antigen receptors (CARs), and have been effective in treating certain cancers such as leukemia.
Application of CAR T-Cell Therapy in Solid tumours
The Car T technology has wowed the field by all but obliterating some patients’ blood cancers, but solid malignancies present new challenges.
Therapies that contains such chimeric antigen receptor (CAR) T cells have been approved for some types of so-called liquid cancers of the blood and bone marrow, large B-cell lymphoma and B-cell acute lymphoblastic leukemia. But the approach has not had as much success for solid tumors.
Serious research into the therapy for brain cancer started almost 20 years ago after cancer biologist WaldemarDebinski, then at Penn State, discovered that the receptor for the immune signaling molecule interleukin 13 (IL-13) was present on glioblastomas, but not on healthy brain tissue. The receptor thus seemed like an excellent target to home in on cancer cells while sparing healthy ones. The CAR spacer domain that spans the immune cells’ membranes and its intracellular co-stimulatory areas, as well as the process used to expand cells outside the body, to boost the T cells’ activity.
CAR T- A Safer Cell Therapy
While managing CAR T-cell therapy toxicity could help keep already-designed treatments on their march to the clinic, many immunotherapy companies are also working to develop a new generation of inherently safer therapies, yet just as efficient. A crucial part of achieving this goal will be improving CAR specificity for target cells. With current treatments, the destruction of normal cells is often an unavoidable side effect when healthy tissue carries the same antigens as tumors; noncancerous B cells, for example, are usually casualties in CD19-targeted therapies.
CAR T delivery is a non-easy factor in the treatment of solid tumors and other unknown forms of tumors. With the non-solid cancers, cells are administered by a blood infusion, and once in circulation, the CAR T can seek out and destroy the rogue cells. For solid tumors, it’s not so simple.
The main drawback of taking cells from a patient and developing them into a cellular immunotherapy product is that the process can take weeks.
Patel tells The Scientist “But for the majority of patients who may not be a candidate or may not have time to wait for such an approach, the idea that there’s off-the-shelf immunotherapy that could potentially as a living drug act against their cancer, I think is a fascinating concept,”
Insulin-producing cells grown in the lab could provide a possible cure for the age-long disease (diabetes).
Type 1 diabetes is an auto¬immune disease that wipes out insulin-producing pancreatic beta cells from the body and raises blood glucose to dangerously high levels. These high levels of Blood sugar level can be even fatal. Patients are being administered insulin and given other medications to maintain blood sugar level. To those who cannot maintain their blood sugar level, they are given beta-cell transplants but to tolerate beta cell transplants; patients have to take immunosuppressive drugs as well.
A report by a research group at Harvard University tells us that they used insulin-producing cells derived from human embryonic stem cells (ESCs) and induced pluripotent stem cells to lower blood glucose levels in mice. Nowadays, many laboratories are getting rapid progress in human stem cell technology to develop those cells that are functionally equivalent to beta-cells and the other pancreatic cell types. Other groups are developing novel biomaterials to encapsulate such cells and protect them against the immune system without the need for immunosuppressant.
Major pharmaceutical companies and life sciences venture capital firms have invested more than $100 million in each of the three most prominent biotechnological industries to bring such treatments into clinical use:
Cambridge
Massachusetts–based companies Semma Therapeutics
Sigilon Therapeutics, and ViaCyte of San Diego
Researchers of UC San Francisco have transformed human stem cells into mature insulin-producing cells for the first time, a breakthrough in the effort to develop a cure for type-1 (T1) Diabetes. Replacing these cells, which are lost in patients with T1 diabetes, has long been a dream of regenerative medicine, but until now scientists had not been able to find out how to produce cells in a lab dish that work as they do in healthy adults.
What is T1 diabetes?
T1 diabetes is an autoimmune disorder that destroys the insulin-producing beta cells of the pancreas, typically in childhood. Without insulin’s ability to regulate glucose levels in the blood, spikes in blood sugar can cause severe organ damage and eventually death. The condition can be managed by taking regular shots of insulin with meals. However, people with type 1 diabetes still often experience serious health consequences like kidney failure, heart disease and stroke. Patients facing life-threatening complications of their condition may be eligible for a pancreas transplant from a deceased donor, but these are rare, and they are supposed to wait a long time.
Researchers have just made a breakthrough that might one day make these technologies obsolete, by transforming human stem cells into functional insulin-producing cells (also known as beta cells) – at least in mice.
“We can now generate insulin-producing cells that look and act a lot like the pancreatic beta cells you and I have in our bodies,” explains one of the team, Matthias Hebrok from the University of California San Francisco (UCSF).
“This is a critical step towards our goal of creating cells that could be transplanted into patients with diabetes.”
Type-1 diabetes is characterized by a loss of insulin due to the immune system destroying cells in the pancreas – hence, type 1 diabetics need to introduce their insulin manually. Although this is a pretty good system, it’s not perfect.
Making insulin-producing cells from stem cells
Diabetes can be cured through an entire pancreas transplant or the transplantation of donor cells that produce insulin, but both of these options are limited because they rely on deceased donors. Scientists had already succeeded in turning stem cells into beta cells, but those cells remained stuck at an early stage in their maturity. That meant they weren’t responsive to blood glucose and weren’t able to secrete insulin in the right way.
Scientists at the University of California San Francisco made a breakthrough in the effort to cure diabetes mellitus type 1.
For the first time, researchers transformed human stem cells into mature insulin-producing cells, which could replace those lost in patients with the autoimmune. There is currently no known way to prevent type-1 (T1) diabetes, which destroys insulin production in the pancreas, limits glucose regulation, and results in high blood sugar levels. The condition can be managed with regular shots of insulin, but people with the disease often experience serious health complications like kidney failure, heart disease, and stroke.
“We can now generate insulin-producing cells that look and act a lot like the pancreatic beta cells you and I have in our bodies,” according to Matthias Hebrok, senior author of a study published last week in the journal Nature Cell Biology.
“This is a critical step toward our goal of creating cells that could be transplanted into patients with diabetes,” Hebrok, director of the UCSF Diabetes Center, said in a statement.
Islets of Langerhans are groupings of cells that contain healthy beta cells, among others. As beta cells develop, they have to separate physically from the pancreas to form these islets.
The team artificially separated the pancreatic stem cells and regrouped them into these islet clusters. When they did this, the cells matured rapidly and become responsive to blood sugar. In fact, the islet clusters developed in ways “never before seen” in a lab. After producing these mature cells, the team transplanted them into mice. Within days, the cells were producing insulin similar to the islets in the mice. While the study has been successful in mice, it still needs to go through more rigorous testing to see if it would work for humans as well. But the research is up-and-coming. “We can now generate insulin-producing cells that look and act a lot like the pancreatic beta cells you and I have in our bodies. This is a critical step towards our goal of creating cells that could be transplanted into patients with diabetes,” He said.
“We’re finally able to move forward on several different fronts that were previously closed to us,” he added. “The possibilities seem endless.”
Basic research keeps elucidating new aspects of beta cells; there seem to be several subtypes, so the gold standard for duplicating the cells is not entirely clear. Today, however, there is “a handful of groups in the world that can generate a cell that looks like a beta cell,” says Hebrok, who currently acts as scientific advisor to Semma and Sigilon, and has previously advised ViaCyte. “Certainly, companies have convinced themselves that what they have achieved is good enough to go into patients.”
The stem cell reprogramming methods that the three companies use to prompt cell differentiation create a mixture of islet cells. Beta cells sit in pancreatic islets of Langerhans alongside other types of endocrine cells. Alpha cells, for example, churn out glucagon, a hormone that stimulates the conversion of glycogen into glucose in the liver and raises blood sugar. Although the companies agree on the positive potential of islet cell mixtures, they take different approaches to developing and differentiating their cells. Semma, which was launched in 2014 to commercialize the Harvard group’s work and counts Novartis among its backers, describes its cells as fully mature, meaning that they are wholly differentiated into beta or other cells before transplantation. “Our cells are virtually indistinguishable from the ones you would isolate from donors,” says Semma chief executive officer BastianoSanna
To get around the donor problem, researchers, including the team at UCSF has been working on nudging stem cells into becoming fully-functional pancreatic beta cells for the last few years. Still, there have been some issues in getting them all the way there.
“The cells we and others were producing were getting stuck at an immature stage where they weren’t able to respond adequately to blood glucose and secrete insulin properly,” Hebrok said.
“It has been a major bottleneck for the field.”
“We’re finally able to move forward on a number of different fronts that were previously closed to us,” Hebrok added. “The possibilities seem endless.”
Regardless of starting cell type, the companies say they are ready to churn out their cells in large numbers. Semma, for example, can make more islet cells in a month than can be isolated from donors in a year in the United States, Sanna says, and the company’s “pristine” cells should perform better than donor islets, which are battered by the aggressive techniques required for their isolation.
As these products, some of which have already entered clinical trials, move toward commercialization, regulatory agencies such as the US Food and Drug Administration (FDA) and the European Medicines Agency have expressed concern about the plasticity of the reprogrammed cells. All three firms subject their cells to rigorous safety testing to ensure that they don’t turn tumorigenic. Before successful trials, companies won’t know the dose of beta cells required for a functional cure, or how long such “cures” will last before needing to be boosted. There’ll be commercial challenges, too: while the companies are investing heavily to develop suitable industrial processes, all acknowledge that no organization has yet manufactured cell therapies in commercial volumes.
Nevertheless, there’s growing confidence throughout the field that these problems will be solved, and soon. “We have the islet cells now,” says Alice Tomei, a biomedical engineer at the University of Miami who directs DRI’s Islet Immuno-engineering Laboratory.
“These stem cell companies are working hard to try to get FDA clearance on the cells.”
Protecting stem cell therapies from the immune system
Whatever the type of cell being used, another major challenge is delivering cells to the patient in a package that guards against immune attack while keeping cells fully functional. Companies are pursuing two main strategies:
Microencapsulation, where cells are immobilized individually or as small clusters, in tiny blobs of a biocompatible gel.
Macroencapsulation, in which greater numbers of cells are put into a much larger, implantable device.
ViaCyte, which recently partnered with Johnson & Johnson, launched its first clinical trial in 2014. The trial involved a micro-encapsulation approach that packaged up the company’s partially differentiated, ESC-derived cells into a flat device called the PEC-Encapsulation. About the size of a Band-Aid, the device is implanted under the skin, where the body forms blood vessels around it. “It has a semipermeable membrane that allows the free flow of oxygen, nutrients, and glucose,” says ViaCyte’s chief executive officer, Paul Laikind. “And even proteins like insulin and glucagon can move back and forth across that membrane, but cells cannot.”
The trial showed that the device was safe, well-tolerated, and protected from the adaptive immune system—and that some cells differentiated into working islet cells. But most cells didn’t engraft effectively because a “foreign body response,” a variant of wound healing, clogged the PEC-Encap’s membrane and prevented vascularization. ViaCyte stopped the trial and partnered with W. L. Gore & Associates, the maker of Gore-Tex, to engineer a new membrane. “With this new membrane,” says Laikind, “we’re not eliminating that foreign body response, but we’re overcoming it in such a way that allows vascularization to take place.” The company expects to resume the trial in the second half of this year, provided it receives the green light from the FDA.
Semma is also developing macro¬-encapsulation methods, including a very thin device that in prototype form is about the size of a silver dollar coin. The device is “deceptively simple, but it allows us to put [in] a fully curative dose of islets,” Sanna says.
Semma is also investigating microencapsulation alternatives. At the same time, the company is advancing toward clinical trials using established transplantation techniques to administer donated cadaver cells to high-risk patients who find it particularly difficult to control their blood glucose levels. These cells are infused via the portal vein into the liver, and patients take immunosuppressive drugs to prevent rejection.
Sigilon is working on its microencapsulation technology. Launched in 2016 on the back of work by the labs of Robert Langer and Daniel Anderson at MIT, the company has created 1.5-millimeter gel-based spheres that can hold between 5,000 and 30,000 cells (Nat Med, 22:306–11, 2016). Each sphere is like a balloon, with the outside chemically modified to provide immune-protection, says Sigilon chief executive officer Rogerio Vivaldi. “The inside of the balloon is full of a gel that creates almost a kind of a matrix net where the cells reside.”
In 2018, shortly after partnering with Eli Lilly, Sigilon and collaborators published research showing that islet cells that were encapsulated in gel spheres and transplanted into macaques remained functional for four months. The company has not disclosed a time frame for a type 1 diabetes trial “but we’re moving pretty quickly,” says chief scientific officer David Moller.
Conclusion
To conclude, all three firms hope to extend their work to treat some of the 400 million people worldwide with type 2 diabetes, many of them eventually benefit from insulin injections. The recent endorsements from big Pharmaceutical underline the real progress in beta-cell transplants, says Aaron Kowalski, a molecular geneticist and chief executive officer at JDRF, a foundation based in New York that has funded research at ViaCyte and academic labs whose work has been tapped by Semma and Sigilon. “These companies all realize that if they don’t do it, somebody else will. It’s hard to predict exactly when, but somebody is going to make this work.”
A lot of viruses that has taken a toll on life, the ebola virus in Africa, The Nipah virus of Nipah and the most recent one corona virus that left china running helter skelter all seemed to have originated from bats. During the course of the virus epidemic in Wuhan where it was first detected, some Chinese researchers in Wuhan examined some patients affected in that area and then took samples of the virus.
They did findings on the genetic sequence of the virus with other viruses that were known. The corona virus surprisingly had a 96% match with the horseshoe bats that are dominant in the southwest of china. The research findings were then published in a study on February, 2020.
A virologist Vineet Menachery from the university of Texas Medical Branch at Galveston though not affiliated to the study said “They’re too close in terms of their pure genetics to say they’re not related, or that they didn’t have a common ancestor.”
Menachery was a reputable virologist and had done other research works. He contributed to the theory that the spread of the corona virus must have been from these bats to humans. And possibly must have had another animal that served as an intermediary for the spread.
This same thing had happened with other forms of corona viruses as noted in the case of SARS (Severe Acute Respiratory Syndrome) an outbreak that took place in 2002-2003 where civets, a mongoose family member were infected with the bat corona virus and spread as humans bought them for food.
Another case was the MERS (Middle East Respiratory Syndrome) outbreak. This one happened in 2012 and was as a result of infected camels from the virus. People who ate undercooked meat of camels and as well drank the raw milk of camels were all affected.
So why is it that there are so many diseases that are spread from bats?
Its no doubt, bats have a lot of viruses that they carry with them. And these viruses in their variety are spread and manifests its tolls on people. Scientist are not sure why this is the case as confirmed by Kevin Olival, a research vice president as EcoHealth Alliance, a non-profit organization based in the U.S. He went further to say that it may have something to do with the family of the viruses carried by the bats. So you know, there are over 130 different families of viruses that bats do carry around.
And then, most bats and humans do come in contact through several means. The millions of populations of bats are ubiquitous to all the continents apart from in antartica. Rebekah of Colorado State university who researched infectious pathogens said “There’s a lot of viruses we’re finding in bats because there’s a lot of bats out there.”
They move about in multitudes and live in colonies of large populations. Some these members live in caves and share caves and trees where there can be a contact between humans and bats. Hence, these viruses can spread from these bats to humans.
Despite their sizes, bats have relatively long lifespans and can live over 30 years.”So there’s a long time for them to be persistently infected with the virus and shed it into the environment,” Kading says. The mode of mechanisms for these viruses are through urine, saliva and feces of bats. The outbreak of Nivah that happened in Bangladesh was linked to the sap of a date palm gotten from some trees that some bats licked and had infested with their urine.
Reading through all these, it is not absurd to wonder why the bats themselves do not get affected by the viruses they carry.
The answer to that question is based on the fact that the bat is the only flying mammal in the world. Their body metabolism and process quite differ from that of normal mammals too. When bats fly, their heart rates rise to about a thousand beats per minute with a temperature rise of about 100 degrees Fahrenheit. Linfa Wang a student of bat viruses at Duke-NUS Medical School in Singapore says that when these signs manifest in other mammals, they are signals that can trigger death. But this is not the same case for other bats. This is a lifestyle for them, every day.
Their system is also capable of producing molecules that other organisms do not have. The molecules carry out repair functions and prevent cell damage. This makes their system a bit irresistible to infections and also make them recalcitrant to viruses and resilient to diseases such as diabetes, cancer and other health conditions.
This is a prove that the manifestation of viruses in mammals is not always as a result of the virus itself, but as a result of the body’s reaction to the presence of such a virus that makes us ill by triggering other chain reactions, as Wang explains.
Olival at EcoHealth explains that these bats have coevolved with these viruses and it is not totally their fault that we humans are infected and affected by these viruses. The actual problem is when the viruses move from their species to other species of mammals which is also fostered by human activity.
Naturally, it would be hard for most animals and mammals to cross paths. But Olivial says that the presence of some activities and availability of exchange platforms made available by humans can allow such interaction to occur. She gave an example using wildlife markets like the one in Wuhan, where a bat could be mixed up with a civet. Who later on come in contact with humans – eg. Butchers who do not observe proper hygiene and protection from animal blood.
“The way that we’re coming into contact with these animals, hunting, selling, and trading them is to a scale that really we haven’t seen before,” he says.
Investigative teams did some in-depth search and they discovered some traces of this virus in 22 stalls and in a garbage truck that was found at Huanan Seafood Market right there in Wuhan, a place known for booming trade for live animals. This discovery led to shutting down the market as it was tied to majority of the cases.
The intermediary animals to this viruses are still a mystery, but it is clear that some of these animals are prone to interact more with humans. This is why when they are infected, the likelihood for human infection is widened. These other infected animals can sneeze, urinate, be cooked as food or even owned as pets.
Bats are not just vectors for viruses, they play an important role in balancing the eco-system. They feed on insects and fruits and are active agents of pollination. In fact, Wang believes that since these bats have successfully coevolved with these viruses, there is every possibility that they can be the agents that can lead to the cure and provision of therapies for these viruses.
Because of the complex nature of wound healing process, an injury on the skin can pose several challenges and are likely pose complications especially when they are acute. They can as well deteriorate from acute to chronic conditions which will require external intervention best understood by a specialist physician to get the area affected by the wound under normalcy.
The complexity of wound healing and research remains an ocean of knowledge that is continuously researched intensely to uncover depths of wound healing techniques and interventions. Hence, this report contains an introduction and details to the use of a new medical innovation called Gcells used primarily for the management of wounds in their different etiology.
In a case where the process of wound healing seemed difficult, Gcells proved great effects an attribute to their design and working protocol. Gcells are conditioned to work with an enriched suspension of progenitor cells that can efficiently aid tissue repair process. In this case report, two subjects were used as donors and acceptors of these micro-grafts.
Introduction
The skin is an outer layer of the body, offering protection to the underlying layers. A wound breaks this layer and inhibits the various functions as well as expose or also break the underlying layer of tissues. Repair processes are inherent and part of homeostatic processes of the body to try to restore the skin back to its normalcy in structure and in function.
The basics for the skins repair mechanism is represented by a cloth and an inflammation where vessels dilate and monocytes activate leading to breakdown of necrotic tissues. This basic process can be inhibited or delayed by a number of varying factors that lead to deteriorative transformation of acute wounds to chronic forms. But if there is no alteration in the repair process, Mesenchymal cells kickstart proliferative process and begin to repair and restructure the affected tissues starting from the base. At the same time epithelial tissues begin to grow around the wound leading to a final step of the healing process. In this final stage, remodeling of the skin structure is primary and then maturation of a scar.
These processes are efficient best in certain conditions which if affected by factors such as cardiovascular ailment, diabetes, bacterial or any other genre of infection, can inhibit these processes.
Hence, it is necessary to understand in details these processes if there is going to be development or innovation for effective healing processes. Just as stated above, during the proliferative phase of wound healing, Mesenchymal cells are the key role players. Their structure includes a Mesenchymal stem cell (MSCs), multi potent in nature and offer supportive, therapeutic and trophic functions. They are also able to release viable trophic, anti-inflammatory cytokines and anti-apoptotic molecules that offer protection during the repair of wounded skin. MSCs also possess subpopulations that are stem-like nature commonly referred to as “side population” (SP) they have been found out to be enriched in over 1000-fold of progenitor cells and multipotent stem cells and as well exist in tissues and tumors. SP exists in a variety of organs and tissues, after an original discovery to be prominent in the bone marrow of a mouse. The organs with SP include the lung, liver, brain, mammary gland and in skeletal muscles.
In other discoveries, it was discovered that they probably may also be isolated in other tissues of the body. This discovery was in an in vitro and in vivo experiment when Dental Pulp Stem Cells (DPSCs) showed capability to differentiate into osteoblasts and built a woven bone by forming an Extracellular Matrix (ECM) secreted by the osteoblasts. The experiment drew results on the both the quality and quantity of the matrix formed by the DPSCs in the in vivo and in vitro experiment using Stem cells and accompanying biomaterials.
Thus proved that dental pulp holds potentialities of therapeutic strength and a rich source of progenitor/autologous cells that can be used to aid healing processes even applicable to regeneration of craniofacial bones.
This is the evidence that supports the working principle of Gcells innovation. Gcell successfully separates this side population with a size of 50 micron. At this cell population, they can form autologous micro-grafts and can either be used alone or alongside biomaterials prepared in a biocomplex ready for use when necessary.
In this case report, two subjects were used as donors and acceptors of these micro-grafts for enhanced healing of complex wounds through autologous micro-grafts using the Gcell.
Clinical case 1
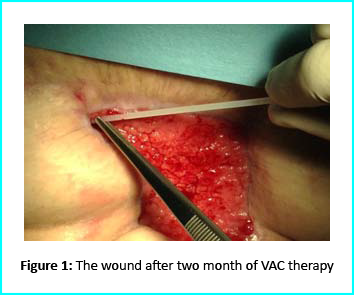
The first case involves a woman at age 50 who does not have any diseases or disorders. She underwent a laparoscopic gastric bypass surgery and was doing well considering parameters of weightloss. Two years later she moved in for abdominoplasty bariatric. Later on, post complications showed preeminence of necrosis which was discovered after first medical examination were about 150 to 200cm2 at the end of the flaps. An initial necrosectomy showed an intense loss of tissue and we furthered to place the wound on VAC therapy and the patient in active participation of this therapy for one week then at home as an outpatient.
As an outpatient, there was improved and progressive wound cleansing while granulated tissues around the base area were cleared.
The VAC therapy after 2 months still left the margins of the wound deteriorated and surrounding areas not in axis with skin surface.
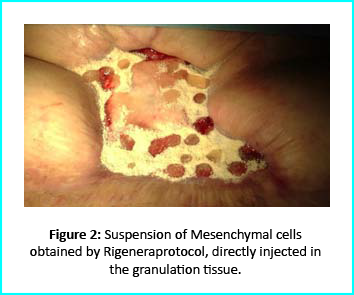
The Gcell protocol kickstarted after consent from the outpatient. We started by collecting a 3 cm2 skin sample from the patient for the purpose of obtaining the cell suspension needed to be injected to the granulation tissue (figure2).
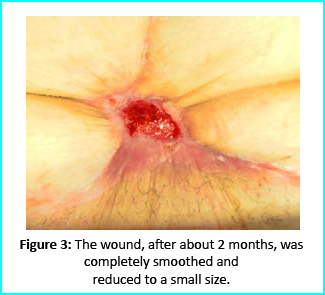
We followed up with conventional wound treatment as in cleaning and replacement with sterile gauze dabbed with Vaseline. The wound area began to improve in both healing progress and general appearance. In two months, the undermined area disappeared as well as leveled to the axis of the skin surface. 2 months later, the wound reduced to a very little scar that is mild and smoothed compared to the initial condition. (figure 3).
A man who suffered liver cirrhosis, hiatal hernia and diabetic as well at the age of 78. Complex surgery was carried out and distal esophagectomy was performed. But hiatal hernia was not decreased into the abdomen, so he was booked up for corrective surgery. During the intervention the adhesions correlated to the previous abdominal operation and led to opening the colon for resection. Some postoperative complications by the appearance of entero-cutaneous fistulas, related to a colonic anastomosis dehiscence. A second intervention was inevitable hence a ileostomy protection and repackaging of colonic anastomosis. We closed the laparotomy using a biological prosthesis. But we met further complications from ascetic failure that needed intensive care hepatology.
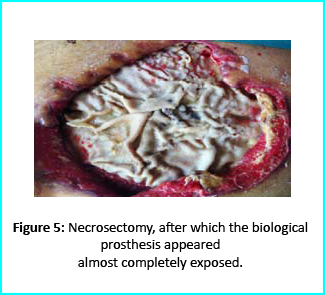
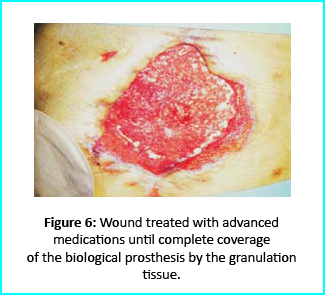
Patient’s condition that included poor liver synthesis had its toll on the healing of the surgical wound. Just as the first case, necrotic tissues grew around the biological prosthesis. We conducted necrosectomy and the biological prosthesis was left half exposed. (Figure 5).
Further treatment of the wound using advanced medication helped cover the biological prosthesis with granulation tissue (figure 6).
Plastic surgeons conducted evaluations on the patient and the choice to do a rotation flap did not seem so appropriate. VAC therapy was used on the wound for about 15 days even though the device wasn’t efficient enough to maintain supposed suction in the presence of ileostomy. We proceeded to treat the patient further with Gcell protocol when wound dimension progressed to about 250cm2. The tissue granulation was of right margin near the ileostomy improved even though it appeared to be undermined.
In summary, Gcell protocol has proved a great level of efficiency in healing and restoration of damaged tissues. This progress is certain to open way for employment in the clinical practice that involves the treatment and management of acute and chronic wounds and in any other field of medicine that will inevitably need an instrument to repair lesion on tissues.
Discussion and conclusion
We made it clear earlier in this document about the efficiency of Gcell protocol in its aid to wound healing especially for wounds that are likely to develop from acute to chronic conditions. The working principle for the Gcell used to obtain the viable progenitor cells used for the micrografts relies on one individual as both the donor and the acceptor. This will help to reduce complications that are related to implants or injected micrografts that are non-autologous. Gcell is flexible and can be used both during in operating rooms as well as in ambulatories. This innovation is vastly spreading and currently used in the fields of oral-maxillo-facial field proven by recent studies even though a greater area of its application widespread and acceptable in plastic surgery, dermatology and orthopedics.
Conclusion of this report brings to clarity in demonstration, an efficient, useful and low-risk innovation in the field of medicine, useful for areas in wound management and healing. However, the viability of the Gcell product still needs to be texted on subjects with different conditions and perspectives. But we assure that this device will prove to be a better therapeutic approach in the field of medicine in improving healing of complex wounds. This confidence lies in the excellent features and working principles of this device in obtaining cell suspension, flexibility, facility for procedure and more importantly, the cost. This will help reduce the use of exorbitantly prices devices for advanced medication.
In summary, apart from introducing an efficient innovation in the medicine. Gcell has the potentialities to offer employment on clinical procedures that will help aid in the management of wounds no matter how the case may be.
One of the amputating surgeries in the field of medicine is a knee replacement. It involves removing the knee joint and replacing it with a modified prosthesis. However, several modifications of this surgery have been introduced into the high-powered world of surgery, including several alternatives for knee replacement. In this article, we are going to review the several modifications and knee replacement alternatives therein.
What is Knee Replacement?
Knee replacement, also known as knee arthroplasty, is a surgical procedure that involves the amputation or cutting out of a knee joint, the bones reams by a doctor, especially due to accidents or joint ailments such as arthritis. When the bone is removed, it is then replaced with a prosthetic device. Knee replacement can be partial, where selected or affected parts of the joints can be removed, such as the medial, lateral, and anterior compartments can also be removed and replaced with a modified prosthetic.
Why Should You Be looking for a Knee Replacement Alternatives?
Due to the dynamics of the human body, what works for the goose may not necessarily work for the gander. Certain post-symptoms of a knee replacement can be unbearable for most patients.
Pain After Knee Replacement.
Due to pain in the knee joint, a lot of patients embark on this old-time surgery to help reduce the pain they feel around their knee. But it is worthy of knowing that a substantial number of these patients still continue to feel pain after this audacious surgery. In a survey done by the government, 40% of patients that underwent knee replacement experienced miniature pain for over 3-4 years, while another 44% still felt some 3-5/10 degree of pain in 3-4 years. So, it is not worthy of looking in the direction of knee replacement alternatives in order to solve knee pain.
Knee Replacement Risks.
There is a risk in everything that we do, business, taking a walk, climbing a hill. Same way, certain risks exist in knee replacement which are:
Patients become more susceptible to heart attack and stroke immediately after knee replacement surgery.
Increased levels of metals in the blood.
Allergic reactions to the prosthetic material.
Possibility of infection.
Reduced activity of the patient as they thrive to become accustomed to the new prosthesis.
Even though social media and digital marketers paint a vivid picture of beautiful seniors riding a bike, continuing in their daily activities and hobbies, but this may not be true for everyone; in a study conducted by the government, there was seldom activity by patients after knee replacement surgery. Another study showed that patients who weren’t running before a knee replacement surgery couldn’t run after the surgery. But there are always two ways to everything; some other patients also showed an increase in physical activity after their surgery.
What are Knee Replacement Alternatives?
Steroid Injections
Steroids are made up of corticosteroids and cortisone. These corticosteroids carry out an anti-inflammatory function to prevent swelling around the knee regions as well as help reduce pain. But they do have a side effect; they destroy cartilage and may not be efficient as they are thought to be. If you are considering this knee replacement alternative, you probably should bear in mind that they do not offer long term remedies. Steroid injections are viable for knee replacement needs caused by arthritis but may proffer short-termed solutions.
Viscosupplementation
Viscosupplementation is also another knee replacement alternative. They are in the form of gels for the knee, also knowns as hyaluronic acid varying across different brands in the market, likes of SynVisc, OrthoVisc, Supartz, and Euflexxa. They are administered to the patient, but a quick question one would ask is if really the shots help. The variations of results all over the web show support both sides of the notion. But one peculiarity of these results is that none says that they are hurtful or damaging as the steroid injections rather that they give a better solution to knee joint arthritis patients. In my own experience, these injections are efficient only when administered a few times, after which they begin to diminish in effects. The first dose may offer relief for some time, but a dose a far-reaching as the sixth dose may not offer any remedial effect at all.
Knee Nerve Ablation
Knee Nerve ablation is another breakthrough in the surgical world. Knee Nerve Ablation involves the use of technology to carry out a process where the specialist probes the nerves around the joint and passes electrical energy that is used to ablate (destroy) them. The work of these nerves is to relay signals from that region of the knee to the brain. So this technique deadens these nerves, and as such, you don’t feel any pain till those nerves grow back. The research on this type of knee replacement alternative is only a handful. Hence, they cannot conclude on the long term results since most of the studies on this new breakthrough are in their early stages.
Orthobiologics
Orthobiologics incorporation around the knee regions helps to enhance the healing of the knew joint or reduce the consequent degradation of orthopedic tissues. Orthobiologics are also knee replacement alternatives and can be gotten from the patient as autologous or a donor as allogeneic. The two primary derivations of orthobiologics are the PRP and the BMC short for Bone Marrow Concentrate. Another derivation that is commonly used is derived from natal tissues as in amniotic or umbilical cord. Just as the nerve ablation, the research on this type of knee replacement is at its early stages.
Platelet Rich Plasma (PRP)
We mentioned PRP earlier while discussing orthobiologics. PRP’s stand for Platelet-rich plasma that can be gotten from the patient. They contain healing factors that allow them to foster cartilage repair as well as reduce inflammation and balance the chemical dynamics of the knee. A lot of studies support the efficiency of PRP as knee replacement alternatives but may not offer much help when the arthritis is severe.
PKA (Percutaneous Knee Arthroplasty)
PKA (Percutaneous Knee Arthroplasty) comes in handy for severe cases of arthritic pain. This procedure involves the injection of rich bone marrow concentrates gotten from the patient or from a donor into the lax ligaments or other affected areas such as damaged meniscus tissues and tendons. This procedure is intricate and uses an ultrasound and fluoroscopy guides as compared to other quick knee shot techniques. Research proves that this method works pretty well, even in extreme cases of knee arthritis. This procedure also produces a lasting effect for about 2-7 years before the need for repetition.
Here you go!! Knee replacement alternatives. You sure would want to consider some of the alternatives; likes of PKA, PRP, and Bone Marrow concentrates that proffers a long-lasting solution.
1American Advanced Medical Corp. (Private Practice), Beverly Hills, CA,
2Western University of Health Sciences, Pomona, CA, USA
Abstract
The prevalence of androgenic alopecia (AGA) increases with age and it affects both men and women. Patients diagnosed with AGA may experience decreased quality of life, depression, and feel self-conscious. There are a variety of therapeutic options ranging from prescription drugs to non-prescription medications. Currently, AGA involves an annual global market revenue of US$4 billion and a growth rate of 1.8%, indicating a growing consumer market. Although natural and synthetic ingredients can promote hair growth and, therefore, be useful to treat AGA, some of them have important adverse effects and unknown mechanisms of action that limit their use and benefits. Biologic factors that include signaling from stem cells, dermal papilla cells, and platelet-rich plasma are some of the current therapeutic agents being studied for hair restoration with milder side effects. However, most of the mechanisms exerted by these factors in hair restoration are still being researched. In this review, we analyze the therapeutic agents that have been used for AGA and emphasize the potential of new therapies based on advances in stem cell technologies and regenerative medicine.
Introduction
The prevalence of androgenic alopecia (AGA) increases with age, and is estimated to affect about 80% of Caucasian men.1 Female AGA, also known as female pattern hair loss, affects 32% of women in the ninth decade of life.2 The consumer market for products that promote hair growth has been increasing dramatically.3 These products promote hair regeneration based on the knowledge about the hair follicle (HF) cycle.4,5 However, in most cases, the mechanisms of action of these products are not well characterized and the results are variable or with undesirable side effects.6 At present, only two treatments for AGA have been approved by the US Food and Drug Administration (FDA): Minoxidil and Finasteride.7–10Although these medications have proved to be effective in some cases, their use is limited by their side effects.11,12 With the emergence of stem cells (SCs), many mechanisms that lead to tissue regeneration have been discovered.13 Hair regeneration has become one of the targets for SC technologies to restore the hair in AGA.14 Several SC factors such as peptides exert essential signals to promote hair regrowth.15,16 Some of these signals stimulate differentiation of SCs to keratinocytes which are important for HF growth.17 Other signals can stimulate dermal papilla cells (DPCs) that promote SC proliferation in the HF.18,19 In this review, we describe HF characteristics and discuss different therapies used currently for AGA and possible novel agents for hair regeneration. These therapies include FDA-approved medications, non-prescription physical or chemical agents, natural ingredients, small molecules, biologic factors, and signals derived from SCs.
HF and SC niche
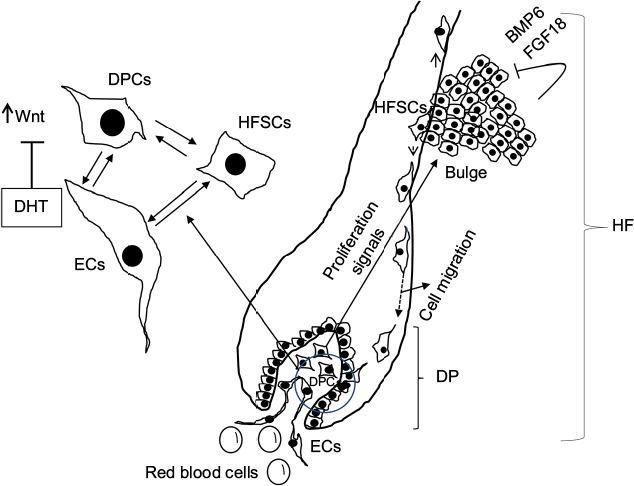
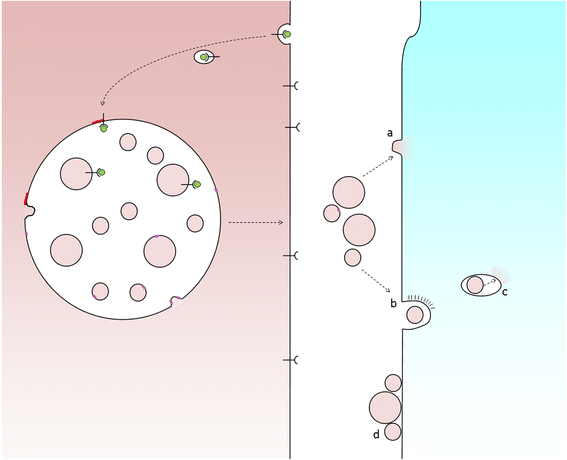
The HF undergoes biologic changes from an actively growing stage (anagen) to a quiescent stage (telogen) with an intermediate remodeling stage (catagen).4 HFSCs are located in the bulge region of the follicle and they interact with mesenchymal SCs (MSCs) located in the dermal papilla (DP).18 These signal exchanges promote activation of some cellular pathways that are essential for DPC growth, function, and survival, such as the activation of Wnt signaling pathway.19–21 Other signals, such as those from endothelial cells (ECs) located at the DP, are also essential for HF maintenance.22 EC dysfunction that impairs adequate blood supply may limits or inhibits hair growth.22 For instance, Minoxidil, a synthetic agent, is able to promote hair growth by increasing blood flow and the production of prostaglandin E2 (PGE2).7 It has been shown that proteins that belong to the transforming growth factor (TGF) superfamily, such as bone morphogenetic proteins (BMPs), also exert signals to maintain the capacity of DPCs to induce HF growing in vivo and in vitro.23 These BMPs may be released by several cells that compose the follicle, including ECs.24–26 ECs may provide signals for BMP receptor activation in DPCs similar to those signals that promote survival of MSCs in human embryoid bodies composed of multipotent cells.24,25 DPCs have been derived from pluripotent SCs in an attempt to study their potential for hair regeneration in vitro and in vivo.27 Together, dermal blood vessels and DPCs orchestrate a suitable microenvironment for the growth and survival of HFSCs.28,29 Interestingly, the expression of Forkhead box C1 regulates the quiescence of HFSCs located in the bulge region (Figure 1).30 HFSCs are quiescent during mid-anagen and maintain this stage until the next hair cycle.29,30 However, during early anagen stage, these cells undergo a short proliferative phase in which they self-renew and produce new hair.30 Therefore, the bulge region constitutes a SC niche that makes multiple signals toward quiescence or proliferation stages.30–34 It is known that fibroblasts and adipocyte signals are able to inhibit the proliferation of HFSCs.34 Additionally, BMP6 and fibroblast growth factor 18 (FGF18) from bulge cells exert inhibitory effects on HFSC proliferation.34 Dihydrotestosterone (DHT) also inhibits HF growth.35 Agents that reduce DHT, such as Finasteride, promote hair regrowth by inhibiting Type II 5a-reductase.8,14,36 In contrast to these inhibitory effects, DPCs located at the base of the HF provide activation signals (Figure 1).18,34 The crosstalk between DPCs and HFSCs leads to inhibition of inhibitory effects with the resultant cell proliferation toward hair regeneration (anagen).30,31,37 With the self-renewal of HFSCs, the outer root sheath (ORS) forms, and signals from DPCs to the bulge cells diminish in a way that the bulge cells start again with their quiescent stage.4,34As mentioned earlier, Forkhead box C1 transcription factor has an important role in maintaining the threshold for HFSC activation.30 The knockdown of these factors in bulge cells reduces the cells’ threshold for proliferation, and the anagen cycle starts more frequently due to promotion of HFSC proliferation in shorter periods of time.30
Figure 1 Diagram of the HF and factors involved in hair regeneration.
Notes: The HF is composed of different cell types including HFSCs, DPCs, and ECs, among others. HFSCs migrate from the bulge area after activation by growth factors released by DPCs. However, BMP6 and FGF18 from the bulge cells exert autocrine inhibitory effects in HFSC proliferation. Once the HFSCs are closer to DPCs and ECs, they differentiate and proliferate during anagen phase, forming new hair. Activation of Wnt signaling is essential for DPCs to release the factors that promote differentiation and proliferation of HFSCs. DHT interferes with this Wnt signaling and, in this way, inhibits hair growth and promotes hair miniaturization. Effective cell–cell interactions between HFSCs, DPCs, and ECs are essential for hair growth.
Prescribed and non-prescription products that promote hair growth and possible mechanisms of action
FDA-approved chemical agents
At present, the only therapeutic agents for AGA approved by the FDA in the USA are Finasteride and Minoxidil.9,10 Minoxidil promotes hair growth by increasing the blood flow and by PGE2 production.7Although Minoxidil is now a non-prescription medication, Finasteride and other drugs require a medical prescription for AGA treatment (Table 1). Dutasteride and Finasteride inhibit 5a-reductase, blocking the conversion of testosterone to DHT.36,38 While Finasteride is a selective inhibitor of type II 5a-reductase, Dutasteride inhibits type I and type II 5a-reductases. These medications have also been used to treat benign prostatic hyperplasia.39
Synthetic prostaglandin analog of PGF2a (originally used to decrease ocular pressure in glaucoma)
Activates prostaglandin receptor
Abbreviation: AGA, androgenic alopecia; PGF2a, prostaglandin F2a.
Natural ingredients
In addition to prescribed medications, some natural ingredients have been used to promote hair growth (Table 2). For example, procyanidin B-2 (found in apples and in several plants) is able to inhibit the translocation of protein kinase C (PKC) in hair epithelial cells.40 PKC isozymes, such as PKC-ßI and -ßII, play an important role in hair cycle progression and inhibiting their translocation can promote hair growth.40 Procyanidin B-3 can promote hair growth by inhibiting TGF-ß1.41 Another group of natural ingredients, such as saw palmetto, alfatradiol, and green tea (Epigallocatechin gallate), have the capacity to inhibit 5a-reductase and block DHT production.42–44 The natural ingredients and their proposed mechanisms of action are summarized in Table 2 (the commercial web page is included, since there are no formal studies about their mechanisms of action).
Table 2
Non-prescription products used for AGA and their proposed mechanisms of action
Natural (flavonoid found in several non-citrus fruits, vegetables, leaves, and grains)
Inhibits PGD2
Abbreviations: AGA, androgenic alopecia; DHT, dihydrotestosterone; ECM, extracellular matrix; FDA, US Food and Drug Administration; PGD2, prostaglandin D2; PKC, protein kinase C; TGF-ß1, transforming growth factor ß1.
Laser therapy
Light amplification by stimulated emission of radiation (LASER) generates electromagnetic radiation which is uniform in polarization, phase, and wavelength.45 Low-level laser therapy (LLLT), also called “cold laser” therapy, since it utilizes lower power densities than those needed to produce heating of tissue. Transdermal LLLT has been used for therapeutic purposes via photobiomodulation.46,47 Several clinical conditions, such as rheumatoid arthritis, mucositis, pain, and other inflammatory diseases, have been treated with these laser devices.48–50 LLLT promotes cell proliferation by stimulating cellular production of adenosine triphosphate and creating a shift in overall cell redox potential toward greater intracellular oxidation.51 The redox state of the cell regulates activation of signaling pathways that ultimately promotes high transcription factor activity and gene expression of factors associated with the cell cycle.52 Physical agents such as lasers have been also used to prevent hair loss in a wavelength range in the red and near infrared (600–1,070 nm).5,47,51,53 Laser therapy emits light that penetrates the scalp and promotes hair growth by increasing the blood flow.54 This increase gives rise to EC proliferation and migration due to upregulation of vascular endothelial growth factor (VEGF) and endothelial nitric oxide synthase.55,56 In addition, the laser energy itself stimulates metabolism in catagen or telogen follicles, resulting in the production of anagen hair.53,54A specific effect of LLLT has been demonstrated to promote proliferation of HFSCs, forcing the hair to start the anagen phase.57
Biologic agents that promote hair growth and their mechanisms of action
SC signaling
Recently, it has been found that SCs release factors that can promote hair growth.16 These factors and their mechanisms of action have been summarized in Table 3. These factors, known as “secretomes”, are able to promote skin regeneration, wound healing, and immunologic modulation, among other effects.58,59 Some of these factors, such as epidermal growth factor (EGF), basic fibroblast growth factor, hepatocyte growth factor (HGF) and HGF activator, VEGF, insulin-like growth factor (IGF), TGF-ß, and platelet-derived growth factor (PDGF), are able to provide signals that promote hair growth.15,60–64 As mentioned before, DPCs provide signals to HFSCs located in the bulge that proliferate and migrate either to the DP or to the epidermis to repopulate the basal layer (Figure 1).32,65 Enhancement in growth factor expression (except for EGF) has been reported when the adipose SCs are cultured in hypoxic conditions.15 Also, SCs increase their self-renewal capacity under these conditions.66–68 Low oxygen concentrations (1%–5%) increase the level of expression of SC factors that include VEGF, basic fibroblast growth factor, IGF binding protein 1 (IGFBP-1), IGF binding protein 2 (IGFBP-2), macrophage colony-stimulating factor (M-CSF), M-CSF receptor (M-CSFR), and PDGF receptor ß (PDGFR-ß).15,69,70 While these groups of factors promote HF growth in intact skin, another group of factors, such as M-CSF, M-CSFR, and interleukin-6, are involved in wound-induced hair neogenesis.71 HGF and HGF activator stimulate DPCs to promote proliferation of epithelial follicular cells.61 Epidermal growth factor promotes cellular migration via the activation of Wnt/ß-catenin signaling.60 VEGF promotes hair growth and increases the follicle size mainly by perifollicular angiogenesis.72 Blocking VEGF activity by neutralizing antibodies reduced the size and growth of the HF.72 PDGF and its receptor (PDGFR-a) are essential for follicular development by promoting upregulation of genes involved in HF differentiation and regulating the anagen phase in HFs.64,73 They are also expressed in neonatal skin cells that surround the HF.73 Monoclonal antibodies to PDGFR-a (APA5) produced failure in hair germ induction, supporting that PDGFR-a and its ligand have an essential role in hair differentiation and development.73 IGF-1 promotes proliferation, survival, and migration of HF cells.69,74 In addition, IGF binding proteins (IGFBPs) also promote hair growth and hair cell survival by regulating IGF-1 effects and its interaction with extracellular matrix proteins in the HF.70 Higher levels of IGF-1 and IGFBPs in beard DPCs suggest that IGF-1 levels are associated with androgens.74 Furthermore, DPCs from non-balding scalps showed significantly higher levels of IGF-1 and IGFBP-6, in contrast to DPCs from balding scalps.74
Table 3
Stem cell factors and small molecules that promote hair growth and their mechanisms of action
Upregulates the genes involved in hair follicle differentiation. Induction and regulation of anagen phase. PDGF and its receptors are essential for follicular development
Small molecules with low molecular weight (<900 Da) and the size of 10-9 m are organic compounds that are able to regulate some biologic processes.75 Some small molecules have been tested for their role in hair growth.76 Synthetic, non-peptidyl small molecules that act as agonists of the hedgehog pathway have the ability to promote follicular cycling in adult mouse skin.76 PGE2 and prostaglandin D2 (PGD2) have also been associated with the hair cycle (Table 3).77 PGD2 is elevated in the scalp of balding men and inhibits hair lengthening via GPR44 receptor.78 Also, it is known that PGE2 and PGF2a promote hair growth, while PGD2inhibits this process.77,79 Prostaglandin analogs of PGF2a have been used originally to decrease ocular pressure in glaucoma with parallel effects in the growth of eyelashes, which suggests a specific effect in HF activation.80 PGD2 receptors are located in the upper and lower ORS region and in the DP, suggesting that these prostaglandins play an important role in hair cycle.81 Molecules such as quercetin are able to inhibit PGD2 and, in this way, promote hair growth.82–84 Antagonists of PGD2 receptor (formally named chemoattractant receptor-homologous expressed in Th2 cells) such as setipiprant have been used to treat allergic diseases such as asthma, but they also have beneficial effects in AGA.85–87 Another small molecule l-ascorbic acid 2-phosphate promotes proliferation of ORS keratinocytes through the secretion of IGF-1 from DPCs via phosphatidylinositol 3-kinase.88 Recently, it has been described that small-molecule inhibitors of Janus kinase–signal transducer and activator of transcription (JAK-STAT) pathway promote hair regrowth in humans.89 Janus kinase inhibitors are currently approved by the FDA for the treatment of some specific diseases such as psoriasis and other autoimmune-mediated diseases.90–94 Also, another group of small molecules such as iron and the amino acid l-Lysine are essential for hair growth (Table 3).95
Cellular therapy
The multipotent SCs in the bulge region of the HF receive signals from DPCs in order to proliferate and survive.27,28,65,84,96 It has been shown that Wnt/ß-catenin signaling is essential for the growth and maintenance of DPCs.19,97 These cells can be isolated and cultured in vitro with media supplemented with 10% fetal bovine serum and FGF-2.37,98 However, they lose versican expression that correlates with decrease in follicle-inducing activity in culture.98 Versican is the most abundant component of HF extracellular matrix.99 Inhibition of glycogen synthase kinase-3 by (2’Z,3’E)-6-bromoindirubin-3′-oxime (BIO) promotes hair growth in mouse vibrissa follicles in culture by activation of Wnt signaling.98 Therefore, the increase of Wnt signaling in DPCs apparently is one of the main factors that promote hair growth.19 DPCs have been also generated from human embryonic SCs that induced HF formation after murine transplantation.27
Platelet-rich plasma
Platelets are anucleate cells generated by fragmentation of megakaryocytes in the bone marrow.100 These cells are actively involved in the hemostatic process after releasing biologically active molecules (cytokines).100–102 Because of the platelets’ higher capacity to produce and release these factors, autologous platelet-rich plasma (PRP) has been used to treat chronic wounds.103 Therefore, PRP can be used as autologous therapy for regenerative purposes, for example, chondrogenic differentiation, wound healing, fat grafting, AGA, alopecia areata, facial scars, and dermal volume augmentation.101,104–108 PRP contains human platelets in a small volume that is five to seven times higher than in normal blood and it has been proven to be beneficial to treat AGA.10,105,109–111 The factors released by these platelets after their activation, such as PDGFs (PDGFaa, PDGFbb, PDGFab), TGF-ß1, TGF-ß2, EGF, VEGF, and FGF, promote proliferation of DPCs and, therefore, may be beneficial for AGA treatment.109,112–114 Clinical experiments indicate that patients with AGA treated with autologous PRP show improved hair count and thickness.109
In search of novel therapies
In this paper, we reviewed and discussed the use of therapeutic agents for hair regeneration and the knowledge to promote the development of new therapies for AGA based on the advances in regenerative medicine. The HF is a complex structure that grows when adequate signaling is provided to the HFSCs. These cells are located in the follicle bulge and receive signals from MSCs located in the dermis that are called DPCs. The secretory phenotype of DPCs is determined by local and circulatory signals or hormones. Recent discoveries have demonstrated that SCs in culture are able to activate DPCs and HFSCs and, in this way, promote hair growth. The study of these cellular signals can provide the necessary knowledge for developing more effective therapeutic agents for the treatment of AGA with minimal side effects. Therefore, advancements in the field of regenerative medicine may generate novel therapeutic alternatives. However, further research and clinical studies are needed to evaluate their efficacy.
Disclosure
The authors report no conflicts of interest in this work.
Blumeyer A, Tosti A, Messenger A, et al; European Dermatology Forum (EDF). Evidence-based (S3) guideline for the treatment of androgenetic alopecia in women and in men. J Dtsch Dermatol Ges. 2011;9(Suppl 6):S1–S57.
Perez-Mora N, Velasco C, Bermüdez F. Oral finasteride presents with sexual-unrelated withdrawal in long-term treated androgenic alopecia in men. Skinmed. 2015;13(3):179–183.
Shapiro J, Kaufman KD. Use of finasteride in the treatment of men with androgenetic alopecia (male pattern hair loss). J Investig Dermatol Symp Proc. 2003;8(1):20–23.
Rossi A, Cantisani C, Melis L, Iorio A, Scali E, Calvieri S. Minoxidil use in dermatology, side effects and recent patents. Recent Pat Inflamm Allergy Drug Discov. 2012;6(2):130–136.
Motofei IG, Rowland DL, Georgescu SR, Mircea T, Baleanu BC, Paunica S. Are hand preference and sexual orientation possible predicting factors for finasteride adverse effects in male androgenic alopecia? Exp Dermatol. 2016;25(7):557–558.
Jain R, De-Eknamkul W. Potential targets in the discovery of new hair growth promoters for androgenic alopecia. Expert Opin Ther Targets. 2014;18(7):787–806.
Park BS, Kim WS, Choi JS, et al. Hair growth stimulated by conditioned medium of adipose-derived stem cells is enhanced by hypoxia : evidence of increased growth factor secretion. Biomed Res. 2010;31(1):27–34.
Zhang P, Kling RE, Ravuri SK, et al. A review of adipocyte lineage cells and dermal papilla cells in hair follicle regeneration. J Tissue Eng. 2014;5:2041731414556850.
Tsai SY, Sennett R, Rezza A, et al. Wnt/ß-catenin signaling in dermal condensates is required for hair follicle formation. Dev Biol. 2014;385(2):179–188.
Choi YS, Zhang Y, Xu M, et al. Distinct functions for Wnt/ß-catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell. 2013;13(6):720–733.
Rendl M, Polak L, Fuchs E. BMP signaling in dermal papilla cells is required for their hair follicle-inductive properties. Genes Dev. 2008;22(4):543–557.
Talavera-Adame D, Wu G, He Y, et al. Endothelial Cells in Co-culture Enhance Embryonic Stem Cell Differentiation to Pancreatic Progenitors and Insulin-Producing Cells through BMP Signaling. Stem Cell Rev. 2011;7(3):532–543.
Talavera-Adame D, Ng TT, Gupta A, Kurtovic S, Wu GD, Dafoe DC. Characterization of microvascular endothelial cells isolated from the dermis of adult mouse tails. Microvascular Res. 2011;82(2):97–104.
Lay K, Kume T, Fuchs E. FOXC1 maintains the hair follicle stem cell niche and governs stem cell quiescence to preserve long-term tissue-regenerating potential. Proc Natl Acad Sci U S A. 2016;113(1): E1506–E1515.
Leirós GJ, Attorresi AI, Balañá ME. Hair follicle stem cell differentiation is inhibited through cross-talk between Wnt/ß-catenin and androgen signalling in dermal papilla cells from patients with androgenetic alopecia. Br J Dermatol. 2012;166(5):1035–1042.
Chacon-Martinez CA, Klose M, Niemann C, Glauche I, Wickström SA. Hair follicle stem cells= cultures reveal self-organizing plasticity of stem cells and their progeny. EMBO J. 2017;36(2):151–164.
Yamana K, Labrie F, Luu-The V. Human type 3 5a-reductase is expressed in peripheral tissues at higher levels than types 1 and 2 and its activity is potently inhibited by finasteride and dutasteride. Horm Mol Biol Clin Investig. 2010;2(3):293–299.
Osada A, Iwabuchi T, Kishimoto J, Hamazaki TS, Okochi H. Long-term culture of mouse vibrissal dermal papilla cells and de novo hair follicle induction. Tissue Eng. 2007;13(5):975–982.
Choi GS, Kim JH, Oh SY, et al. Safety and tolerability of the dual 5-alpha reductase inhibitor dutasteride in the treatment of androgenetic alopecia. Ann Dermatol. 2016;28(4):444–450.
Kamimura A, Takahashi T. Procyanidin B-3, isolated from barley and identified as a hair-growth stimulant, has the potential to counteract inhibitory regulation by TGF-beta1. Exp Dermatol. 2002;11(6):532–541.
Ulbricht C, Basch E, Bent S, et al. Evidence-based systematic review of saw palmetto by the Natural Standard Research Collaboration. J Soc Integr Oncol. 2006;4(4):170–186.
Rondanelli M, Perna S, Peroni G, Guido D. A bibliometric study of scientific literature in Scopus on botanicals for treatment of androgenetic alopecia. J Cosmet Dermatol. 2015;15(2):120–130.
Blume-Peytavi U, Kunte C, Krisp A, Garcia Bartels N, Ellwanger U, Hoffmann R. [Comparison of the efficacy and safety of topical minoxidil and topical alfatradiol in the treatment of androgenetic alopecia in women.] J Dtsch Dermatol Ges. 2007;5(5):391–395. German.
Bjordal JM, Couppé C, Chow RT, Tunér J, Ljunggren EA. A systematic review of low level laser therapy with location-specific doses for pain from chronic joint disorders. The Aust J Physiother. 2003;49(2):107–116.
Brosseau L, Welch V, Wells G, et al. Low level laser therapy (classes I, II and III) in the treatment of rheumatoid arthritis. Cochrane Database Syst Rev. 2005;19(2):CD002049.
Jimenez JJ, Wikramanayake TC, Bergfeld W, et al. Efficacy and safety of a low-level laser device in the treatment of male and female pattern hair loss: a multicenter, randomized, sham device-controlled, double-blind study. Am J Clin Dermat. 2014;15(2):115–127.
Tuby H, Maltz L, Oron U. Modulations of VEGF and iNOS in the rat heart by low level laser therapy are associated with cardioprotection and enhanced angiogenesis. Lasers Surg Med. 2006;38(7):682–688.
Avci P, Gupta GK, Clark J, Wikonkal N, Hamblin MR. Low-level laser (light) therapy (LLLT) for treatment of hair loss. Laser Surg Med. 2015;46(2):144–151.
Rodrigues C, de Assis AM, Moura DJ, et al. New therapy of skin repair combining adipose-derived mesenchymal stem cells with sodium carboxymethylcellulose scaffold in a pre-clinical rat model. PloS One. 2014;9(5):e96241.
Zhang H, Nan W, Wang S, et al. Epidermal growth factor promotes proliferation and migration of follicular outer root sheath cells via Wnt/ß-catenin signaling. Cell Physiol Biochem. 2016;39(1):360–370.
Lee YR, Yamazaki M, Mitsui S, Tsuboi R, Ogawa H. Hepatocyte growth factor (HGF) activator expressed in hair follicles is involved in in vitro HGF-dependent hair follicle elongation. J Dermatol Sci. 2001;25(2):156–163.
Ohyama M, Terunuma A, Tock CL, et al. Characterization and isolation of stem cell – enriched human hair follicle bulge cells. J Clin Invest. 2006;116(1):249–260.
Grayson WL, Zhao F, Bunnell B, Ma T. Hypoxia enhances proliferation and tissue formation of human mesenchymal stem cells. Biochem Biophys Res Commun. 2007;358(3):948–953.
Ren H, Cao Y, Zhao Q, et al. Proliferation and differentiation of bone marrow stromal cells under hypoxic conditions. Biochem Biophys Res Commun. 2006;347(1):12–21.
Osaka N, Takahashi T, Murakami S, et al. ASK1-dependent recruitment and activation of macrophages induce hair growth in skin wounds. J Cell Biol. 2007;176(7):903–909.
Sasaki S, Hozumi Y, Kondo S. Influence of prostaglandin F2alpha and its analogues on hair regrowth and follicular melanogenesis in a murine model. Exp Dermatol. 2005;14(5):323–328.
Zhang Q, Major MB, Takanashi S, et al. Small-molecule synergist of the Wnt/beta-catenin signaling pathway. Proc Natl Acad Sci U S A. 2007;104(18):7444–7448.
Fong P, Tong HH, Ng KH, Lao CK, Chong CI, Chao CM. In silicoprediction of prostaglandin D2 synthase inhibitors from herbal constituents for the treatment of hair loss. J Ethnopharmacol. 2015;175:470–480.
Weng Z, Zhang B, Asadi S, et al. Quercetin is more effective than cromolyn in blocking human mast cell cytokine release and inhibits contact dermatitis and photosensitivity in humans. PloS One. 2012;7(3):e33805.
Norman P. Update on the status of DP2 receptor antagonists; from proof of concept through clinical failures to promising new drugs. Expert Opin Investig Drugs. 2014;23(1):55–66.
Pettipher R. The roles of the prostaglandin D(2) receptors DP(1) and CRTH2 in promoting allergic responses. Br J Pharmacol. 2008;153(Suppl 1):S191–S199.
Garza LA, Liu Y, Yang Z, et al. Prostaglandin D2 inhibits hair growth and is elevated in bald scalp of men with androgenetic alopecia. Sci Transl Med. 2012;4(126):126ra34.
Kwack MH, Shin SH, Kim SR, et al. I-Ascorbic acid 2-phosphate promotes elongation of hair shafts via the secretion of insulin-like growth factor-1 from dermal papilla cells through phosphatidylinositol 3-kinase. Br J Dermatol. 2009;160(6):1157–1162.
Quintás-cardama A, Vaddi K, Liu P, et al. INCB018424 : therapeutic implications for the treatment of myeloproliferative neoplasms Preclinical characterization of the selective JAK1 / 2 inhibitor INCB018424 : therapeutic implications for the treatment of myeloproliferative neoplasms. Blood. 2014;115(15):3109–3117.
Bao L, Zhang H, Chan LS. The involvement of the JAK-STAT signaling pathway in chronic inflammatory skin disease atopic dermatitis. JAKSTAT. 2013;2(3):1–8.
Oshima H, Rochat A, Kedzia C, Kobayashi K, Barrandon Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell. 2001;104(2):233–245.
Yamauchi K, Kurosaka A. Inhibition of glycogen synthase kinase-3 enhances the expression of alkaline phosphatase and insulin-like growth factor-1 in human primary dermal papilla cell culture and maintains mouse hair bulbs in organ culture. Arch Dermatol Res. 2009;301(5):357–365.
Sotoodehnejadnematalahi F, Burke B. Structure, function and regulation of versican: the most abundant type of proteoglycan in the extracellular matrix. Acta Med Iran. 2013;51(11):740–750.
Xu XR, Zhang D, Oswald BE, et al. Platelets are versatile cells: new discoveries in hemostasis, thrombosis, immune responses, tumor metastasis and beyond. Crit Rev Clin Lab Sci. 2016;53(6):409–430.
Gentile P, Garcovich S, Bielli A, Scioli MG, Orlandi A, Cervelli V. The Effect of platelet-rich plasma in hair regrowth: a randomized placebo-controlled trial. Stem Cells Transl Med. 2015;4(11):1317–1323.
Lynch MD, Bashir S. Applications of platelet-rich plasma in dermatology: a critical appraisal of the literature. J Dermatolog Treat. 2016;27(3):285–289.
Trink A, Sorbellini E, Bezzola P, et al. A randomized, double-blind, placebo- and active-controlled, half-head study to evaluate the effects of platelet-rich plasma on alopecia areata. Br J Dermatol. 2013;169(3):690–694.
Oh Y, Kim BJ, Kim MN. Depressed facial scars successfully treated with autologous platelet-rich plasma and light-emitting diode phototherapy at 830 nm. Ann Dermatol. 2014;26(3):417–418.
Alves R, Grimalt R. Randomized placebo-controlled, double-blind, half-head study to assess the efficacy of platelet-rich plasma on the treatment of androgenetic alopecia. Dermatol Surg. 2016;42(4):491–497.
Cho JW, Kim SA, Lee KS. Platelet-rich plasma induces increased expression of G1 cell cycle regulators, type I collagen, and matrix metalloproteinase-1 in human skin fibroblasts. Int J Mol Med. 2012;29(1):32–36.
Leo MS, Kumar AS, Kirit R, Konathan R, Sivamani RK. Systematic review of the use of platelet-rich plasma in aesthetic dermatology. J Cosmet Dermatol. 2015;14(4):315–323.
Exosomes are extracellular vesicles first described as such 30 years ago and since implicated in cell–cell communication and the transmission of disease states, and explored as a means of drug discovery. Yet fundamental questions about their biology remain unanswered. Here I explore what exosomes are, highlight the difficulties in studying them and explain the current definition and some of the outstanding issues in exosome biology.
What is the current definition of an exosome?
That is a very good question. Since the original description of exosomes over 30 years ago, the term has been loosely used for various forms of extracellular vesicle, muddying the field and contributing to the scepticism with which the research has sometimes been met. Exosomes are best defined as extracellular vesicles that are released from cells upon fusion of an intermediate endocytic compartment, the multivesicular body (MVB), with the plasma membrane. This liberates intraluminal vesicles (ILVs) into the extracellular milieu and the vesicles thereby released are what we know as exosomes (Fig. 1).
Exosomes correspond to intraluminal vesicles of multivesicular bodies. A transmission electron micrograph of an Epstein–Barr virus-transformed B cell displaying newly expelled exosomes at the plasma membrane. Multivesicular bodies (MVB) can be seen which can deliver content to lysosomes for degradation or can fuse with the cell surface to release intraluminal vesicles as exosomes, indicated by the arrows at the top of the picture
There are other types of microvesicle, including apoptotic bodies and ectosomes, which are derived from cells undergoing apoptosis and plasma membrane shedding, respectively. Although apoptotic bodies, ectosomes and exosomes are all roughly the same size (typically 40–100 nm) and all also contain ‘gulps’ of cytosol, they are different species of vesicles and understanding differences between them is of paramount importance but has too often been overlooked.
How were exosomes first recognized as distinct entities?
The presence of membranous vesicles outside cells was first recognized 50 years ago, although these were originally assumed to be waste products released via shedding of the plasma membrane. The recognition of what we now call exosomes didn’t come until 1983, from studies on the loss of transferrin during the maturation of reticulocytes into erythrocytes [1]. These studies showed, by following transferrin-gold conjugates through the endocytic system, that ILVs generated in MVBs can be released to the extracellular space through fusion with the plasma membrane [2], although it was not until 1987 that the term ‘exosome’ was coined for them [3].
Even then, however, these extracellular vesicles were largely ignored, forgotten or, again, dismissed as a means of cellular waste disposal. It is only in the past decade that interest in exosomes has exploded, with a nearly tenfold increase in publications in as many years (115 in 2006, 1010 in 2015).
Why this explosion of interest?
For at least three reasons. First, they are thought to provide a means of intercellular communication and of transmission of macromolecules between cells. Second, in the past decade, exosomes have been attributed roles in the spread of proteins, lipids, mRNA, miRNA and DNA and as contributing factors in the development of several diseases. And third, they have been proposed to be useful vectors for drugs because they are composed of cell membranes, rather than synthetic polymers, and as such are better tolerated by the host. In fact, some of the earliest exosome research indicated that they can carry the MHC–peptide complexes that are recognized by T lymphocytes [4] and that secretion of such exosomes could promote antitumour immune responses in mice in vivo [5]. Exosome therapies are now being explored in anti-cancer clinical trials and recent reports claim taxol-filled exosomes can be used to treat cancers in mice at 50-fold lower doses than conventional treatments, with the additional benefit that exosomes do not invoke an immune response [6].
Yet despite 20 years of research, the very basics of exosome biology are in their infancy and we know little of the part they play in normal cellular physiology.
So do we know how they are generated?
Yes and no. We do know that they are made as ILVs; but first of all, not all ILVs finish up as exosomes, and second, the mechanism of their generation in endosomes is not fully understood. Most conventional membrane budding processes deform membrane from an organelle into the cytoplasm but in ILV formation the membrane buds away from the cytoplasm and into the endosome. This unconventional budding process is not limited to ILV generation but also takes place during enveloped virus budding from the cytosol and during cytokinesis [7], and it requires specialised machinery.
ILVs (and thus exosomes) can be generated at the endosomal limiting membrane by at least two mechanisms, one of which depends on the ESCRT machinery (ESCRT stands for endosomal sorting complexes required for transport) whereas the other is ESCRT-independent (Fig. 2).
ILVs are generated by invagination of the endosomal membrane and have three possible fates. Inset: intraluminal vesicles (ILV) are formed by invagination of the endosomal membrane by either ESCRT-dependent or ESCRT-independent mechanisms. Matured endosomes accumulate ILVs within their lumen and have three distinct fates. They may deliver content that contributes to the biogenesis of specialized lysosome-related organelles (for example, melanosomes, Weibel-Palade bodies, azurophilic granules), they may fuse with lysosomes or they may fuse with the plasma membrane where released ILVs are now termed ‘exosomes’
The ESCRT machinery consists of a set of cytosolic protein complexes that are recruited to endosomes by membrane proteins that have been tagged, usually with ubiquitin on their cytosolic domains. The ubiquitin tag is recognized by the first of the ESCRT complexes, ESCRT-0, which is thus recruited to the endosomal membrane and passes ubiquitinated cargos to ESCRT-I, one of whose components, Tsg101, also recognizes ubiquitin. The recruitment of the ESCRT machinery acts to both cluster the ubiquitinated cargo proteins on the endosome and induce curvature of the endosomal membrane to form ILVs.
But ILVs are still able to form in the absence of ESCRTs [8], so other means of generating ILVs must exist, although the mechanisms for their generation are less clear. Generation of these ESCRT-independent ILVs requires the tetraspanin CD63—a protein abundant on ILVs but with unclear function [9]—and may be facilitated by cone-shaped bending properties of lipids such as ceramide [10].
If not all ILVs become exosomes, what determines the fate of an ILV?
The destiny of ILVs is directed by the fate of the MVB they reside in. Confusingly, in addition to different types of ILVs, there are also different types of MVBs [11] and what regulates the fate of these endosomes is another interesting question. MVBs have several potential fates (Fig. 2) and can either fuse with lysosomes (where contents are degraded and recycled), fuse with the plasma membrane (where ILVs are released as exosomes), as I have already mentioned, or contribute to the generation of specialised organelles, such as melanosomes (in melanocytes), Weibel-Palade bodies (endothelial cells), azurophilic granules (in neutrophils) and secretory granules (in mast cells). The levels of cholesterol on MVBs appear to play a part in regulating their fate, cholesterol-rich MVBs being directed to the plasma membrane for exosome release, while cholesterol-poor MVBs are targeted to the lysosome [12].
But what regulates the balance between exosome release and alternative fates of ILVs remains engimatic.
What about differences between cells: do all cells release exosomes?
Well, not all cells have an endomembrane system, so no. But most mammalian cells contain endomembranes and generate ILVs within MVBs, though remarkably little is known about exosome release in most cell types.
Some cells—for example, the B cells, dendritic cells and mast cells of the immune system—appear to release exosomes constitutively; in fact, most of the data we have on exosomes comes from immune cells. As well as releasing exosomes constitutively, these cells may also be stimulated to secrete exosomes by cellular interactions. For example, murine dendritic cells, which are specialized to activate T lymphocytes, secrete higher levels of exosomes upon interaction with antigen-specific CD4+ T lymphocytes [13]. In fact, lymphocyte interactions generally can be accompanied by exosome release; human T cells (including primary T cells from blood, T cell clones and Jurkat cell lines) release exosomes upon activation of their antigen receptors [14] and B cells release more exosomes upon engagement with antigen-specific CD4+ T cells [15].
Other cell types can be pushed to secrete exosomes by means of calcium ionophores or other stimuli[16, 17], but the extent of physiological exosome secretion in non-immune cells is largely unknown.
What happens when exosomes reach an acceptor cell?
We don’t know exactly. Exosomes that transfer membrane proteins or luminal content to the acceptor cell may be engulfed whole or the exosome membrane may fuse directly with the host plasma membrane (Fig. 3). Alternatively, exosomes may not need to be taken up by cells to elicit a physiological response: follicular dendritic cells, for example, carry on their cell surface exosomes that bear MHC–peptide complexes and other proteins that they do not express and are thereby enabled to activate immune cells with which they interact [18].
Exosome uptake by recipient cells. Fusion of MVBs with the cell surface releases ILVs as exosomes. In order for exosomes to elicit a response from recipient cells they might either fuse with plasma membrane (a) or be taken up whole via endocytosis (b), following which the exosome must be delivered to the cytosol, for example, via a back-fusion event (c). Alternatively, exosomes may attach to the surface of recipient cells to elicit a signalling response (d)
For intercellular transmission, various mechanisms of phagocytosis and endocytosis of extracellular vesicles have been described and which mechanism operates may depend upon vesicle size, which may in turn depend upon the cargo carried by the vesicle. In order for material to be released to an acceptor cell, exosomes must fuse with the host cell and this takes place via either direct fusion with the plasma membrane or a ‘back-fusion’ step from within a host endocytic organelle after the exosome has been engulfed. The process of back-fusion is not entirely clear, although it appears to require the unconventional lipid LBPA and protein Alix [19] (and is exploited by anthrax toxin lethal factor to escape from endosomes to the cytosol [20]).
Whether exosomes fuse with target cells or act via interactions with cell-surface proteins, or both, is another fundamental cell biology question that will need to be addressed if we are to understand the functions of exosomes.
So what are the consequences of all this information transfer? What biological functions have been established for exosomes?
There are many proposed functions for exosomes, the best-established being in immune responses. Exosomes isolated from B lymphocytes and bearing MHC class II molecules were shown in early experiments [4] to activate T lymphocytes in vitro, suggesting that they were communicating with the T lymphocytes in just the way that the parent B cells did. I have already mentioned later work by the same group, who showed that exosomes derived from dendritic cells, which are specialized to activate T cells in the initiation of immune responses, could promote antitumour immune responses in mice [5], exciting interest in the possibility of practical applications.
Or, as with follicular dendritic cells, exosome-associated MHC II can be found on the surface of cell types that neither express MHC II nor secrete exosomes, indicating that exosomes are delivered from one cell type to another [18].
However, exosomes may have roles other than in immune responses as several non-immune cells secrete exosomes. The only physiological role so far reported for non-immune cells is in keratinocyte-derived exosomes, which have been shown to modulate melanin synthesis by increasing the expression and activity of proteins within the melanosomes that modulate skin pigmentation [21].
How exactly would exosomes from one cell influence the expression and activity of proteins in an acceptor cell?
Exosomes transfer not only protein and lipids but mRNA and microRNA into acceptor cells and these RNAs have been shown in experiments in vitro to have functional effects in recipient cells. For example, exosomes from mice can be transferred to human cells and mRNA can be translated into mouse protein [22]. Similarly, microRNAs—double-stranded RNA fragments that can regulate specific sets of mRNA (and so protein levels)—can act functionally in acceptor cells. The mode of action of exosomes has been a focus of special interest in cancer biology. Exosomes from breast cancer cell lines, for example, have been shown to be enriched for miRNAs relative to nontumorigenic breast cell lines and exposure of normal cells to exosomes derived from breast cancer cell lines increased both cell survival and proliferation, accompanied by loss of expression of some tumour-suppressor proteins [23]. Exosome levels are elevated in the serum of some cancer patients versus controls. However, whether these vesicles are exosomes or other forms of extracellular vesicle, or a mix, is unclear—I have already mentioned this persistent problem in exosome research.
So exosomes can also contribute to disease?
Yes indeed. As exosomes provide a means of intercellular communication, they may also act as vehicles for ‘bad’ communication or spread. As well as miRNAs in the case of cancer, exosomes have been shown to contain numerous disease-associated cargos—for example, neurodegenerative-associated peptides, such as Aß [24] (in Alzheimer’s disease), tau [25] (in numerous neurodegenerative diseases), prions [26] (in transmissible spongiform encephalopathies), alpha-synuclein [27] (in synucleinopathies, including Parkinson’s disease) and superoxide dismutase 1 [28] (in amyotrophic lateral sclerosis). Exosomes have thus been suggested to be propagators of neurodegenerative protein spread, although some cargos are easier to envisage than others.
Of the neurodegenerative-associated proteins, only some are integral membrane proteins, that is, proteins inserted into lipid bilayers, rather than cytosolic. Sorting of proteins into ILVs (and thus exosomes) is easier to envisage for membrane proteins, where tags such as ubiquitin regulate where they end up. So far, the presence of both Aß [29] and PrPc [26] has in fact been shown in ILVs, though this has not been demonstrated for other membrane proteins, such as alpha-synuclein and tau.
The mechanism whereby cytosolic proteins may be sorted to ILVs/exosomes, however, is not clear. In order for cytosolic proteins to become concentrated in ILVs, they would require positive incorporation and sorting, possibly by membrane-associated components on endosomes. All we can say is that there is evidence that this does in fact happen; cytosolic factors such as miRNAs are enriched in exosomes relative to cytosol, indicating that sorting must occur whereby certain miRNAs are concentrated and others are not [30].
The means by which disease-associated factors spread between cells remains poorly understood and exosomes would provide a means for such transmission. The presence of exosomal proteins, such as Alix, in association with Alzheimer’s senile plaques strengthens the circumstantial case for exosomes as a mediator in such spread. The hope is that having a means to regulate exosome release and spread may be useful in combatting some of these diseases but much more basic biology needs to be established before then.
Now I’m confused—what determines what exosomes contain?
Exosomes will contain whatever is sorted into them during their formation (as ILVs). For membrane proteins, this usually occurs through ubiquitination, which acts as a substrate for recruitment of the ESCRT machinery and subsequent generation of ESCRT-dependent ILVs.
The mechanisms that concentrate cytosolic factors are currently unknown. Although it seems clear that miRNAs, for example, are enriched relative to the amount in their parent cells, and are not randomly incorporated into exosomes, it is not clear how some are enriched more than others. There are currently a few hypotheses for miRNA sorting, including sorting via sumoylated heterogeneous nuclear ribonucleoproteins [31] or by a miRNA-induced silencing complex (miRISC) [32].
Because of the difficulties in separating exosomes from other extracellular vesicles, it is likely that some cargos reported to be enriched in ‘exosomes’ may in fact be contained in contaminant vesicles that are not exosomes. While many researchers are very stringent about applying the labels ‘exosomes’ and ‘extracellular vesicles’ correctly, others unfortunately are not. In addition, as I have said before, cytosolic proteins are likely to be found in exosome preparations because the exosome lumen is made of cytosol.
So how exactly can you be sure that a given extracellular vesicle is an exosome and not something else?
This is an interesting question that has a complex answer. Ideally, an intracellular compartment is identified by a specific biological marker, as, for example, in the case of the Golgi, nucleus or mitochondria, all of which carry proteins not found, or found at much lower levels, elsewhere.
One problem is that ILVs, and thus exosomes, represent an intermediate compartment of an intermediate. MVBs are not static organelles but rather undergo continuous maturation, in the course of which they gain and lose proteins. There will never be an exclusive marker for exosomes because any cargo on the ILV/exosome membrane must first be on the limiting membrane of the endosome and anything found inside must first come from the cytosol. A cargo may be concentrated on ILVs/exosomes but it will also be elsewhere. CD63 could be thought of as a pseudo-marker for exosomes. ILVs and exosomes are enriched in several such tetraspanins and my colleagues and I have show that CD63 is required for ESCRT-independent ILV formation [9]. Alix also appears to be concentrated in ILVs/exosomes [33], as does Tsg101, a component of ESCRT-I, which has been used as a marker of exosomes in numerous studies [33,34], although the presence of Tsg101 in ILVs or exosomes does not fit with conventional models of ILV formation. Although Tsg101 is involved in ESCRT-dependent ILV formation, as mentioned earlier, it, along with other ESCRT components, should disassociate from the endosomal membrane prior to an ILV pinching off the endosomal membrane to allow it to participate in further events [35]. Exactly when ESCRT-I components ‘fall off’ the membrane is unknown but it is conventionally thought to be prior to ILV formation, so Tsg101 should remain cytosolic and available for subsequent rounds of ILV formation. It is possible that some Tsg101 may be ‘swallowed’ into the forming ILV lumen, but levels should be negligible.
So are you saying there is no reliable marker for endosomes?
There may not be—not a single reliable one. Ultimately, perhaps the best method of defining exosomes biochemically may be through a combination of markers, including tetraspanins, Alix and others, with a concomitant exclusion of resident plasma membrane proteins. Although ILVs/exosomes will by their nature contain some plasma membrane proteins and the plasma membrane will contain some ILV/exosomal proteins, it should be possible to define relative levels and/or enrichment of proteins of exosomes that distinguish them from other microvesicles. Cargos such as MHC II from B cells and other cell type-specific antigens may also help to distinguish exosomes from other forms of extracellular vesicle. Common exosomal cargos include tetraspanins (CD63, CD81, CD9), antigen presentation molecules (MHC I and MHC II) and others (Alix, flotillin-1). An online database exists [36] where proteins, lipids and RNA are catalogued from published and unpublished exosomal studies.
If they are so hard to characterize reliably, how are exosomes isolated and studied?
Exosomes are rarely imaged by conventional methods as they are too small to be resolved by fluorescence microscopy and their release may be a rare event. A few studies have imaged exosome release occurring in cell cultures by various electron microscopic techniques but, more commonly, exosomes are pooled from cellular supernatant or animal fluids. Traditionally, they have been isolated by differential centrifugation from culture medium whereby larger contaminants are first excluded by pelleting out through increasing speeds of centrifugation before exosomes, small extracellular vesicles and even protein aggregates are pelleted at very high speeds (~100,000?×?g) [37]. These preparations therefore represent an enrichment rather than a purification. Enriched preparations are commonly analysed by biochemistry, mass spectrometry or electron microscopy. Electron microscopy of isolated fractions as ‘whole mounts’ make it possible to immuno-label vesicles, with the limitation that isolated preparations do not provide the same internal controls as labelling sections of cells. Remarkably little attention has been paid to the characterization of exosomes, although efforts are being made to repair this omission with guidelines and criteria for defining groups of extracellular vesicles [38].
What would you say are the most important issues in exosome research?
Without doubt the single most important issue is actually understanding the biological significance of these structures. With so little known about their basic physiological functions, it may seem hard to understand how exosomes have been implicated in the pathogenesis of so many disparate disease states. Fundamental questions remain about exosome generation, fate and normal function but, ultimately, in order to understand exosomes, one must first understand ILVs, a fact that is too often overlooked. Meanwhile, it is important that publications on exosomes give a careful and explicit account of the criteria used for distinguishing them from other extracellular vesicles to avoid confusing the field and encouraging scepticism.
References
Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol. 1983;97(2):329–39.
Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101(3):942–8.
Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–20.
Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, et al. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med. 1998;4(5):594–600.
Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, Klyachko NL, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2015;12(3):655–64.
McDonald B, Martin-Serrano J. No strings attached: the ESCRT machinery in viral budding and cytokinesis. J Cell Sci. 2009;122(Pt 13):2167–77.
Stuffers S, Sem Wegner C, Stenmark H, Brech A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic. 2009;10(7):925–37.
Edgar JR, Eden ER, Futter CE. Hrs- and CD63-dependent competing mechanisms make different sized endosomal intraluminal vesicles. Traffic. 2014;15(2):197–211.
Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–7.
White IJ, Bailey LM, Aghakhani MR, Moss SE, Futter CE. EGF stimulates annexin 1-dependent inward vesiculation in a multivesicular endosome subpopulation. EMBO J. 2006;25(1):1–12.
Mobius W, Ohno-Iwashita Y, van Donselaar EG, Oorschot VM, Shimada Y, Fujimoto T, et al. Immunoelectron microscopic localization of cholesterol using biotinylated and non-cytolytic perfringolysin O. J Histochem Cytochem. 2002;50(1):43–55.
Buschow SI, Nolte-’t Hoen EN, van Niel G, Pols MS, ten Broeke T, Lauwen M, et al. MHC II in dendritic cells is targeted to lysosomes or T cell-induced exosomes via distinct multivesicular body pathways. Traffic. 2009;10(10):1528–42.
Blanchard N, Lankar D, Faure F, Regnault A, Dumont C, Raposo G, et al. TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/zeta complex. J Immunol. 2002;168(7):3235–41.
Muntasell A, Berger AC, Roche PA. T cell-induced secretion of MHC class II-peptide complexes on B cell exosomes. EMBO J. 2007;26(19):4263–72.
Savina A, Furlan M, Vidal M, Colombo MI. Exosome release is regulated by a calcium-dependent mechanism in K562 cells. J Biol Chem. 2003;278(22):20083–90.
Guo BB, Bellingham SA, Hill AF. Stimulating the release of exosomes increases the intercellular transfer of prions. J Biol Chem. 2016;291(10):5128–37.
Denzer K, van Eijk M, Kleijmeer MJ, Jakobson E, de Groot C, Geuze HJ. Follicular dendritic cells carry MHC class II-expressing microvesicles at their surface. J Immunol. 2000;165(3):1259–65.
Bissig C, Gruenberg J. ALIX and the multivesicular endosome: ALIX in Wonderland. Trends Cell Biol. 2014;24(1):19–25.
Abrami L, Lindsay M, Parton RG, Leppla SH, van der Goot FG. Membrane insertion of anthrax protective antigen and cytoplasmic delivery of lethal factor occur at different stages of the endocytic pathway. J Cell Biol. 2004;166(5):645–51.
Lo Cicero A, Delevoye C, Gilles-Marsens F, Loew D, Dingli F, Guere C, et al. Exosomes released by keratinocytes modulate melanocyte pigmentation. Nat Commun. 2015;6:7506.
Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–9.
Melo SA, Sugimoto H, O’Connell JT, Kato N, Villanueva A, Vidal A, et al. Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell. 2014;26(5):707–21.
Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, et al. Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci U S A. 2006;103(30):11172–7.
Saman S, Kim W, Raya M, Visnick Y, Miro S, Saman S, et al. Exosome-associated tau is secreted in tauopathy models and is selectively phosphorylated in cerebrospinal fluid in early Alzheimer disease. J Biol Chem. 2012;287(6):3842–9.
Fevrier B, Vilette D, Archer F, Loew D, Faigle W, Vidal M, et al. Cells release prions in association with exosomes. Proc Natl Acad Sci U S A. 2004;101(26):9683–8.
Emmanouilidou E, Melachroinou K, Roumeliotis T, Garbis SD, Ntzouni M, Margaritis LH, et al. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J Neurosci. 2010;30(20):6838–51.
Gomes C, Keller S, Altevogt P, Costa J. Evidence for secretion of Cu, Zn superoxide dismutase via exosomes from a cell model of amyotrophic lateral sclerosis. Neurosci Lett. 2007;428(1):43–6.
Guduric-Fuchs J, O’Connor A, Camp B, O’Neill CL, Medina RJ, Simpson DA. Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genomics. 2012;13:357.
Edgar JR, Willen K, Gouras GK, Futter CE. ESCRTs regulate amyloid precursor protein sorting in multivesicular bodies and intracellular amyloid-beta accumulation. J Cell Sci. 2015;128(14):2520–8.
Villarroya-Beltri C, Gutierrez-Vazquez C, Sanchez-Cabo F, Perez-Hernandez D, Vazquez J, Martin-Cofreces N, et al. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat Commun. 2013;4:2980.
Fabian MR, Sonenberg N. The mechanics of miRNA-mediated gene silencing: a look under the hood of miRISC. Nat Struct Mol Biol. 2012;19(6):586–93.
Bobrie A, Colombo M, Krumeich S, Raposo G, Thery C. Diverse subpopulations of vesicles secreted by different intracellular mechanisms are present in exosome preparations obtained by differential ultracentrifugation. J Extracell Vesicles. 2012;1:18397.
Thery C, Boussac M, Veron P, Ricciardi-Castagnoli P, Raposo G, Garin J, et al. Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J Immunol. 2001;166(12):7309–18.
Thery C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protocols Cell Biol. 2006;Chapter 3:Unit 3 22.
Lotvall J, Hill AF, Hochberg F, Buzas EI, Di Vizio D, Gardiner C, et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International Society for Extracellular Vesicles. J Extracell Vesicles. 2014;3:26913.
Acknowledgements
The author wishes to thank Scottie Robinson, Paul Luzio and Paul Manna for critical reading of this article.
Competing interests
The author declares that he has no competing interests.
Author information
Affiliations
Cambridge Institute for Medical Research, University of Cambridge, Hills Road, Cambridge, CB2 0XY, UK






")